引言:脚下世界的惊人发现
当人类仰望星空,追寻外星生命的蛛丝马迹时,一个远比火星或欧罗巴更近的"外星世界"正静静地躺在我们脚下。在地球表面之下数公里的深处,在永恒的黑暗与极端的压力中,存在着一个庞大得令人难以置信的生命王国——深部生物圈。
这个地下世界的规模几乎难以想象:据深碳观测站(Deep Carbon Observatory)十年研究的估算,地下生物圈的总体积约为20至23亿立方公里,几乎是地球上所有海洋体积的两倍。其中的微生物总质量在150至230亿吨之间,数百倍于地球上所有人类的总重量。
更令人震撼的是,这些深部生命中约有70%属于细菌和古菌两大生命域,占地球整个生物圈生物量的15%。换句话说,每当你走在地球上,你脚下踩着的可能是一个比地表生命更为庞大的生命网络。
这些微生物生活在极端环境中:温度可高达122°C,压力相当于数百个大气压,没有任何阳光,营养物质极其匮乏。然而,它们不仅存活下来,还进化出了令人惊叹的生存策略。有些微生物的代谢速度慢到令人咋舌,一个细胞分裂可能需要数百年甚至数千年;有些则以岩石中的矿物为食,将放射性衰变产生的能量转化为生命所需的化学能。
深部生物圈的发现,不仅彻底改变了我们对地球生命分布的认知,更为理解生命起源、外星生命存在的可能性,以及地球本身如何成为一个宜居星球,提供了全新的视角。
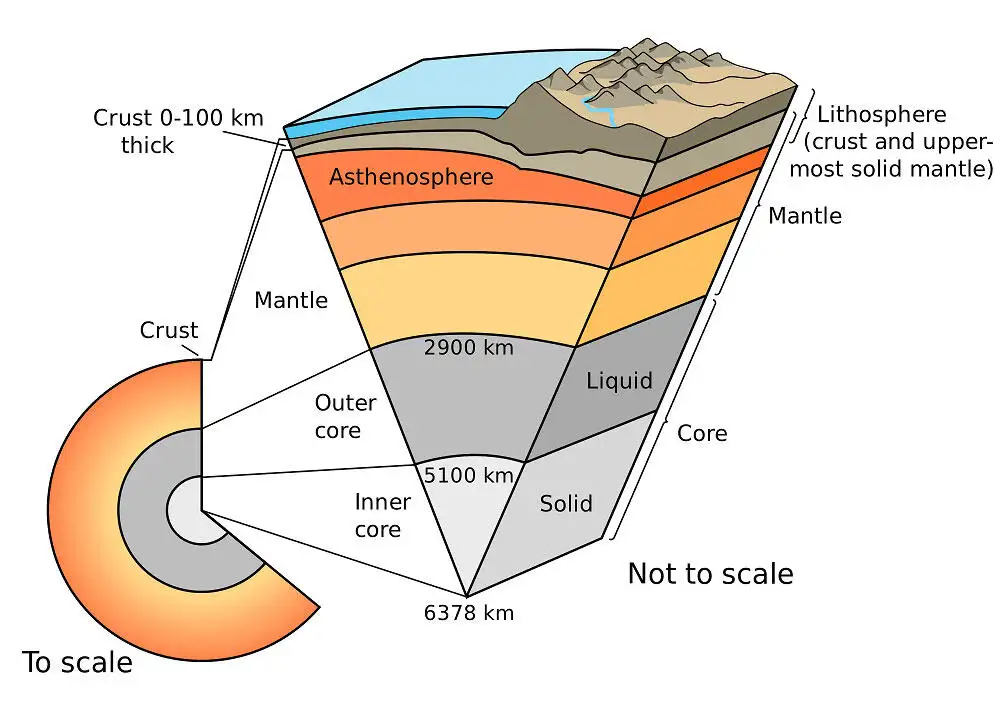
深部生物圈的科学发现历程
早期的怀疑与探索
深部生命存在的可能性,在很长一段时间内都被科学界视为无稽之谈。传统观点认为,生命需要阳光、适中的温度和充足的营养物质——这些条件在地壳深处几乎都不存在。
20世纪20年代,苏联科学家提出了"深部生物圈"的概念,但当时缺乏足够的证据支持。直到20世纪80年代,美国地质调查局的科学家在深层地下水样本中发现了活细菌,这一发现才真正引起了科学界的关注。
然而,真正的转折点出现在1990年代。当时,科学家们开始系统性地研究深层矿井和海洋钻探样本。在南非的威特沃特斯兰德金矿中,研究人员在地下3.5公里深处的水样中发现了大量微生物。这些微生物生活在封闭了数百万年的地下水中,完全独立于地表生态系统。
深碳观测站的十年征程
2011年,深碳观测站(Deep Carbon Observatory)启动,这是一个汇聚了来自52个国家1200多名科学家的国际合作项目。其目标是在十年内系统性地探索地球深部的碳循环和生命分布。
这个项目的发现彻底颠覆了我们对地下生命的认知。研究团队从全球各地的深矿井、深海钻探点和地下钻孔中收集了数千个样本,深度从地下几米延伸到地下5公里以上。
研究结果显示,地下生物圈的规模远超此前所有的估计。在不同深度的样本中,科学家们发现了种类繁多的微生物,从普通的细菌到极端耐热的古菌,从以氢为食的化能自养生物到能够利用辐射能的奇特生命形式。

深部生命的生存策略
化学合成:不依赖阳光的生命
地表生命几乎完全依赖光合作用——植物和藻类利用阳光将二氧化碳和水转化为有机物质。然而,在地下深处的永恒黑暗中,这种能量来源根本不存在。
深部微生物进化出了完全不同的生存策略:化学合成。它们利用无机化学反应释放的能量来维持生命活动。最常见的能量来源是氢气(H₂),它可以与二氧化碳(CO₂)反应生成甲烷(CH₄),释放出可被微生物利用的能量。
这一过程被称为"产甲烷作用",由一类称为产甲烷菌的古菌完成。这些微生物不呼吸氧气——事实上,氧气对它们来说是有毒的——而是利用氢气作为电子供体,将二氧化碳还原为甲烷。
另一种重要的化学合成途径是硫酸盐还原。某些细菌能够将硫酸盐(SO₄²⁻)还原为硫化氢(H₂S),同时氧化有机物或氢气来获取能量。这种代谢方式在深层地下水中非常普遍。
氢气的来源:生命的燃料
既然氢气是深部生命的关键燃料,那么这些氢气从何而来?科学家们发现了多种天然产氢机制:
蛇纹石化反应:当海水或地下水与地幔岩石(主要是橄榄岩)发生反应时,会产生氢气。这种反应在海洋中脊的热液喷口附近尤为活跃,也是著名的"失落之城"(Lost City)热液喷口系统的能量基础。
放射性分解:地下岩石中的铀、钍等放射性元素衰变时释放的辐射可以分解水分子,产生氢气和氧气。这种机制在富含放射性矿物的岩石中尤其重要。
机械化学作用:地震活动或板块运动造成的岩石破碎和摩擦也可以产生氢气。
这些天然产氢机制为深部微生物提供了持续的能量来源,使它们能够在几乎没有任何外部能量输入的环境中生存数百万年。
极慢的代谢:时间的另一种尺度
深部生物圈中最令人惊叹的发现之一,是微生物代谢的极端缓慢。在地表环境中,细菌通常每隔几十分钟或几小时分裂一次。但在地下深处,情况完全不同。
研究显示,某些深部微生物的分裂周期可能长达数百年甚至数千年。这意味着一个细胞从诞生到分裂可能需要经历整个人类文明的历史。
这种极慢的代谢速度反映了地下环境的极端贫瘠。营养物质极其有限,能量获取极其困难,微生物不得不将几乎所有的代谢活动降至最低水平。
然而,这种"慢生活"也有其优势。以如此缓慢的速度消耗资源,微生物可以在一个封闭的环境中存活数百万年甚至数十亿年。2012年,科学家在南太平洋海底以下近600米的沉积物中发现了已经存活了1亿年的微生物。当被带回实验室并提供营养物质后,这些微生物迅速恢复活跃,开始生长和繁殖。
单一物种生态系统:孤独的生存者
2018年,科学家在南非一座金矿地下2.8公里处发现了一个堪称奇迹的生命群落。在这个深处的裂缝水中,他们发现了一个完全由单一物种构成的生态系统。
这种微生物被命名为Candidatus Desulforudis audaxviator(拉丁语意为"大胆的旅行者"),是一种硫酸盐还原细菌。在这个封闭的地下环境中,它独自完成了所有生态功能:从岩石中获取能量、固定碳元素、回收死细胞中的营养物质。
这是科学家首次发现如此简单、完全由单一物种构成的生态系统。通常,生态系统需要多个物种相互配合,形成复杂的食物网。但Desulforudis audaxviator打破了这个规则,它独立支撑起了一个完整的生态系统。
这种微生物的基因组包含约2000个基因,远少于大多数自由生活的细菌,但足以让它在极端环境中自给自足。它不依赖任何来自地表的有机物质,而是完全依靠地球内部提供的能量和碳源。
南非金矿:深部生命的天然实验室
威特沃特斯兰德的地下世界
南非的威特沃特斯兰德盆地是地球上最古老、最大的金矿区之一。这里的金矿可以追溯到约30亿年前,而金矿所在的地层更是形成于约27亿年前。
这些金矿为研究深部生命提供了得天独厚的条件。首先,矿井延伸极深,最深处超过4公里,温度可达60°C以上。其次,岩石中的裂缝含有古老的地下水,这些水可能与地表隔绝了数百万年。
2006年,普林斯顿大学的科学家团队在Mponeng金矿地下约2.8公里处取得了重大发现。在一个岩石裂缝中,他们发现了被封闭了约2000万年的地下水。水中生活着大量细菌,其中大多数属于一种新发现的硫酸盐还原菌。
这个发现的重要意义在于:这些微生物完全独立于太阳能量。它们不依赖光合作用产生的有机物质,而是利用地下岩石中的化学反应来获取能量。
不依赖太阳的生命
地球上的绝大多数生命都直接或间接依赖太阳。植物通过光合作用将太阳能转化为化学能,动物通过食用植物或其他动物获取能量。即使是深海中的生物,大多也依赖从上层海洋沉降下来的有机物质。
但Mponeng金矿中的微生物打破了这一规则。它们利用的能量来自地下岩石中的放射性元素衰变。铀、钍等元素衰变释放的辐射分解水分子,产生氢气。这些微生物利用氢气与岩石中的硫酸盐反应,获取生存所需的能量。
这种代谢方式被称为"辐射营养"(radiotrophy),是地球上最奇特的生命策略之一。它意味着这些微生物可以在完全隔绝于地表的环境中生存——只要有岩石和地下水存在,它们就能持续繁衍。
这个发现对天体生物学具有重大意义。它暗示着在火星或其他行星的地下深处,即使没有阳光,也可能存在类似的生命形式。

海洋深部生物圈:海床之下的隐秘世界
深海沉积物中的生命
海洋覆盖了地球表面的约71%,而海床之下同样是一个巨大的生命栖息地。深部生物圈在海洋沉积物中尤为丰富,从浅海大陆架到深海平原,微生物几乎无处不在。
海洋钻探项目(Ocean Drilling Program)和国际大洋发现计划(International Ocean Discovery Program)在过去几十年中钻取了大量海底沉积物样本。研究表明,微生物可以生活在海床以下超过2.5公里的深处。
这些微生物面临的环境极其严酷:温度随深度增加而上升,压力巨大,营养物质极其匮乏。沉积物中的有机物质在沉降过程中已经被上层微生物消耗殆尽,到达深层的有机碳少之又少。
尽管如此,科学家们仍在深海沉积物中发现了数量惊人的微生物。据估计,海洋沉积物中的微生物总生物量可达地球生物圈总生物量的十分之一。
一亿年的休眠与复苏
2020年,日本海洋研究开发机构(JAMSTEC)的科学家取得了一项突破性发现。他们在南太平洋环流区的海底沉积物中发现了已经"沉睡"超过一亿年的微生物。
这些微生物生活在海床以下约75米处的沉积物中。该区域是地球上最为贫瘠的海域之一,表层海水中的营养物质极少,沉降到海底的有机物质更是微乎其微。沉积物中的氧气含量却出奇地高,因为没有什么微生物消耗它。
研究团队从沉积物样本中提取了微生物,并在实验室中向它们提供营养物质。令人惊讶的是,这些微生物迅速苏醒,开始生长和繁殖。短短65天内,微生物数量增加了四个数量级,从每立方厘米约1000个细胞增长到超过100万个。
这一发现表明,某些微生物可以在极端贫瘠的环境中存活数千万年甚至更长时间,只需极少的能量维持基本的细胞功能。当条件改善时,它们能够迅速恢复活跃。
这种"休眠-复苏"的能力对理解生命的极限具有重要意义。它暗示着在其他行星或卫星的地下环境中,即使环境曾经适宜生命存在但后来变得极端恶劣,微生物仍可能以休眠状态存活,等待条件改善。
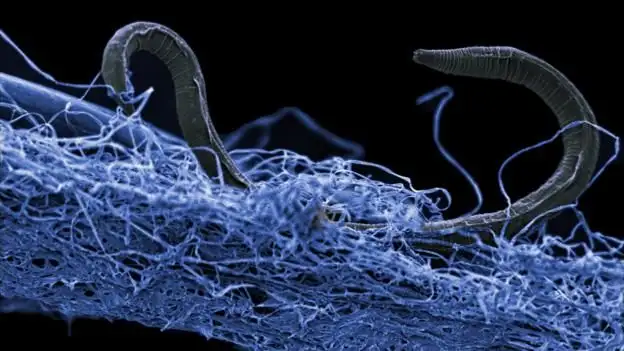
热液喷口:深部生命的绿洲
失落之城的白色塔楼
2000年12月,科学家在大西洋中脊发现了一个独特的热液喷口系统,因其幽灵般白色碳酸钙塔楼而被命名为"失落之城"(Lost City)。
与典型的"黑烟囱"热液喷口不同,失落之城的水温相对较低(约40-90°C),喷出的液体呈碱性,富含氢气和甲烷。这些流体是由海水与地幔岩石发生蛇纹石化反应产生的。
失落之城为研究深部生命提供了一个天然的窗口。这里的热液流体直接来自地球深部,其中的微生物群落展示了生命如何在地球内部的热量和化学能驱动下繁衍生息。
失落之城的微生物群落以氢气氧化细菌为主,它们利用热液中的氢气与海水中的氧气反应获取能量。这些细菌又为更复杂的生物如管虫、贝类和甲壳类动物提供了食物来源。
巨型管虫:共生关系的典范
在深海热液喷口周围,生活着一种令人惊叹的生物——巨型管虫(Riftia pachyptila)。这些蠕虫可以长到2米以上,却没有嘴巴、消化道或肛门。它们如何获取营养?
答案是共生关系。巨型管虫体内生活着大量的硫氧化细菌。管虫将富含硫化物的热液输送到体内的细菌处,细菌氧化硫化物产生能量,并将部分合成的有机物质提供给宿主。
这种共生关系是深部生物圈中常见的生存策略。在缺乏阳光和外部食物来源的环境中,生物之间的合作变得至关重要。管虫提供物理保护和化学原料,细菌提供营养物质,双方互利互惠。

极端生命的物理极限
温度的边界
生命能够承受的最高温度是多少?这个问题对理解深部生物圈至关重要,因为地下深处的温度随深度增加而上升。
目前已知能够生长繁殖的最高温度记录是约122°C,由一种名为Geogemma barossii的古菌保持。这种微生物发现于深海热液喷口,能够在高温高压环境下利用铁离子进行呼吸。
理论上,生命的温度极限可能更高。蛋白质和核酸在高温下会变性,但某些极端微生物进化出了特殊的分子机制来稳定这些生物大分子。例如,它们的热休克蛋白能够帮助其他蛋白质保持折叠状态,而特殊的脂质结构则能够稳定细胞膜。
深碳观测站的科学家估计,在地下约5公里深处(大陆地壳)或海底以下约10公里深处(海洋地壳),温度可能达到生命的极限。但这个界限在哪里,仍然是一个开放的问题。
压力的挑战
地下深处的压力同样是一个巨大的挑战。在地下5公里处,压力相当于约1500个大气压。大多数地表生物在这样的压力下会被压碎。
然而,深部微生物进化出了独特的适应机制。它们的细胞膜含有特殊的脂质,能够在高压下保持流动性。此外,它们还能产生"压力保护分子",如特定氨基酸和糖类,帮助稳定细胞内的蛋白质。
有趣的是,某些微生物不仅能够承受高压,甚至需要高压才能正常生长。这些"嗜压生物"在常压下反而生长缓慢甚至停止生长。这表明深部生物圈的微生物已经完全适应了它们所处的极端环境。
能量的最低门槛
生命维持需要多少能量?这是理解深部生物圈的另一个关键问题。在地表环境中,微生物通常能够获取充足的能量和营养物质。但在地下深处,能量来源极其有限。
研究发现,深部微生物的能量需求低得惊人。它们可以将大部分能量用于维持基本的细胞功能,而非生长和繁殖。某些微生物可能几十年甚至几百年才分裂一次。
这种极低能量代谢的实现,依赖于高度优化的细胞机制。深部微生物通常具有较小的基因组,减少了复制和维持DNA所需的能量。它们的蛋白质合成效率更高,代谢途径更加精简。某些微生物甚至可以进入一种近乎"休眠"的状态,将代谢活动降至接近零。
托马斯·戈尔德的远见
“深部热生物圈"假说
1992年,天体物理学家托马斯·戈尔德(Thomas Gold)提出了一个大胆的假说:地球深部存在一个庞大的生物圈,其中的生命数量可能超过地表所有生命的总和。
戈尔德认为,地下深处可能存在着一个独立的"深部热生物圈”,其中的生命不依赖太阳能,而是利用地球内部的热量和化学能。他将这个地下世界称为"我们这个星球上最大的生态系统"。
当时,戈尔德的观点被许多科学家视为异想天开。然而,随着深部生物圈研究的发展,他的许多预测被证实是正确的。地下微生物的数量和生物量确实远超此前的估计,而化学合成也是这些微生物的主要能量来源。
戈尔德的假说甚至更进一步:他提出地下的碳氢化合物(如石油和天然气)可能并非来自远古生物遗骸的转化,而是由地球深部的无机过程生成。这个"无机成因说"仍然存在争议,但它促使科学家重新思考地球碳循环的复杂性。
从异端到主流
深部生物圈的研究经历了从被忽视到被接受的过程。最初,许多科学家怀疑地下发现的微生物可能只是钻探过程中带入的地表污染。随着采样技术和分析方法的发展,研究者能够证明这些微生物确实来自地下深处,而且与地表微生物有明显的遗传差异。
今天,深部生物圈已经成为地球科学和微生物学研究的一个重要领域。深碳观测站的十年研究彻底改变了我们对地下生命的认知,也验证了戈尔德等人几十年前的远见。

对天体生物学的启示
火星的地下生命?
深部生物圈的发现对寻找地外生命具有重大意义。如果地球上的微生物能够在地下深处、独立于阳光的环境中生存,那么其他行星的地下是否也可能存在类似的生命?
火星是目前最有希望的地外生命候选地。虽然火星表面干燥、寒冷、暴露在致命的辐射下,但其地下可能存在液态水层。火星地壳中的氢气和其他还原性气体可能为化学合成微生物提供能量来源。
事实上,地球上发现的某些深部微生物,如Desulforudis audaxviator,可以作为火星生命的模型。这些微生物展示了生命如何在一个封闭、黑暗、贫瘠的环境中完全依靠地球内部提供的能量生存。
欧罗巴和恩克拉多斯
木星的卫星欧罗巴和土星的卫星恩克拉多斯被认为是太阳系中最有可能存在地外生命的天体之一。这两颗卫星的冰层之下被认为存在液态海洋,深度可能达到数十公里。
在这些海洋的底部,可能存在类似地球失落之城的热液喷口系统。海水与岩石的反应产生氢气和其他还原性物质,为化学合成微生物提供能量来源。
NASA即将发射的欧罗巴快船任务(Europa Clipper)将详细探测这颗卫星的冰壳和地下海洋。虽然这次任务不会直接寻找生命,但它将为未来的生命探测任务奠定基础。
深部生物圈的研究为这些天体生物学探索提供了重要的理论和实践指导。它告诉我们,生命可以在我们曾经认为不可能的环境中生存,只要存在液态水和化学能来源。
深部生物圈与地球演化
地球引擎的参与者
深部微生物不仅是被动的地下居民,它们还积极参与地球的化学和地质过程。通过代谢活动,这些微生物影响着岩石的风化、矿物的形成和元素的循环。
例如,某些深部微生物能够将可溶性金属离子转化为不溶性矿物,从而影响矿石的分布。硫酸盐还原菌产生的硫化氢可以与金属离子反应,形成硫化物矿床。这可能是地球上某些金矿和铜矿形成的重要机制。
微生物还参与着碳循环的关键环节。产甲烷菌产生的甲烷是一种重要的温室气体,它会通过地质活动释放到大气中,影响地球的气候。据估计,地球上大部分的甲烷可能来自深部生物圈的微生物活动。
生命与地球的协同进化
深部生物圈的存在表明,生命与地球之间的关系远比我们想象的更加紧密。生命不仅存在于地球的表面,还渗透到地壳深处,成为地球系统不可分割的一部分。
从地球历史的早期开始,微生物可能就已经在地壳中定居。这些早期生命可能对地球表面的环境产生了深远的影响,例如通过产生氧气改变大气成分,或通过促进矿物风化加速土壤形成。
一些科学家甚至提出,深部生物圈可能是地球生命的起源地。在地球形成的早期,地表环境极其不稳定,频繁的陨石撞击和剧烈的火山活动可能不利于生命的起源和早期演化。而地下深处则提供了一个相对稳定、有液态水和化学能来源的环境,可能更适合生命的诞生。
这个"深部起源假说"虽然尚未得到证实,但它为我们理解生命起源提供了新的视角。如果生命确实起源于地下深处,那么深部生物圈中的微生物可能保留着地球上最古老的生命形式。

探索深部生物圈的技术挑战
采样的难题
研究深部生物圈面临着重重技术挑战。首先是采样问题:如何在不引入污染的情况下,获取地下数公里深处的微生物样本?
传统的钻探方法使用水和泥浆作为润滑剂,这些流体很容易将地表微生物带入地下。为了解决这个问题,科学家们开发了专门的"洁净钻探"技术,使用无菌流体并严格控制钻探过程。
另一种方法是利用现有的深矿井和钻孔。南非的金矿、加拿大的镍矿和德国的钾盐矿都成为研究深部生物圈的天然实验室。科学家可以直接从矿井的裂缝中采集地下水样本,避免了钻探污染的问题。
深海钻探的突破
在海洋环境中,研究深部生物圈更加困难。深海钻探需要专门的钻探船,如著名的JOIDES Resolution号。这艘船可以在数千米深的海域钻取海底以下的沉积物和岩石样本。
2023年春季,国际大洋发现计划在大西洋中脊的亚特兰蒂斯海岭进行了历史性的钻探。科学家们钻入了海底以下超过1.2公里的地幔岩石,创造了深海钻探的新纪录。
这次钻探不仅获取了大量珍贵的岩石和流体样本,还直接触及了活跃的热液系统。科学家们在钻孔深处发现了高浓度的氢气,这为研究深部微生物的能量来源提供了直接证据。
分子生物学的革命
现代分子生物学技术极大地推动了深部生物圈的研究。通过环境DNA测序,科学家可以鉴定样本中存在的所有微生物种类,而不需要在实验室中培养它们。
大多数深部微生物无法在实验室条件下培养,因为它们的生长需求过于特殊。环境DNA测序绕过了这个限制,让研究者能够了解这些微生物的遗传特征和代谢潜力。
此外,单细胞基因组学技术可以对单个微生物细胞进行基因测序,揭示其完整的基因组信息。这对于研究低生物量、低多样性的深部微生物群落尤为重要。

深部生物圈的未来研究方向
探索生命的极限
深部生物圈研究的一个核心问题是:生命的极限在哪里?科学家们仍在探索微生物能够承受的最高温度、最大压力和最低能量供应。
未来的研究将深入更深的地下,寻找可能存在的生命边界。一些科学家甚至提出,生命可能存在于地下10公里甚至更深的处,只要有液态水和化学能来源。
全球碳循环
深部生物圈在全球碳循环中的作用仍然是一个开放的问题。地下微生物产生的甲烷有多少最终进入大气?深部碳循环与地表碳循环如何相互作用?这些问题对于理解地球气候系统至关重要。
深碳观测站的研究表明,地球深部的碳含量远超此前估计。地下微生物在碳的转化和迁移中扮演着重要角色,但具体机制仍有待阐明。
生物技术应用
深部微生物可能具有独特的生物技术潜力。它们产生的酶可能在极端条件下(高温、高压、低pH)保持活性,适用于工业生产。某些深部微生物可能能够降解污染物或生产有价值的化学品。
此外,研究深部微生物的极端适应机制,可能为开发新型抗生素、工业催化剂或环境修复技术提供灵感。
结语:重新认识我们的星球
深部生物圈的发现彻底改变了我们对地球生命的认知。它告诉我们,生命不仅存在于地球的表面,还渗透到地壳深处,形成一个庞大而隐秘的生命网络。
这个地下世界远比我们想象的更加丰富多彩。从南非金矿深处独立于太阳的细菌群落,到南太平洋海底一亿年沉睡后复苏的微生物,从失落之城白色塔楼周围的化学合成群落,到威特沃特斯兰德独自支撑生态系统的"大胆旅行者"——这些发现展示了生命的顽强与创造力。
深部生物圈的研究也深刻影响了我们对外星生命的思考。如果地球上的微生物可以在地下深处、黑暗中、贫瘠的环境中生存,那么其他行星的地下是否也可能孕育着生命?火星、欧罗巴、恩克拉多斯——这些曾经被视为死寂的天体,现在成为了寻找地外生命的热门目标。
当我们凝视脚下的土地时,我们看到的不仅是岩石和土壤,而是一个庞大的生命王国。这个王国或许比地表世界更加古老、更加神秘、也更加重要。它是地球生命史的关键篇章,也是我们理解生命本质和宇宙中生命分布的关键线索。
深部生物圈的探索才刚刚开始。在未来的岁月里,科学家们将继续向下钻探、深入分析,揭开这个地下世界的更多秘密。每一次发现,都可能颠覆我们对生命的认知;每一个新物种,都可能讲述着生命起源和演化的古老故事。
在我们脚下,一个比海洋更深、比天空更神秘的世界,正在等待着被探索。
参考文献
-
Onstott, T. C. et al. (2006). “The Origin and Age of Biogeochemical Trends in Deep Fracture Water of the Witwatersrand Basin, South Africa.” Geomicrobiology Journal, 23(6), 369-414.
-
Chivian, D. et al. (2008). “Environmental Genomics Reveals a Single-Species Ecosystem Deep Within Earth.” Science, 322(5899), 275-278.
-
Kallmeyer, J. et al. (2012). “Global Distribution of Microbial Abundance and Biomass in Subseafloor Sediment.” Proceedings of the National Academy of Sciences, 109(40), 16213-16216.
-
Magnabosco, C. et al. (2018). “The Depth and Temperatures of the Subsurface Biosphere.” Geobiology, 16(3), 259-268.
-
Heberling, C. et al. (2010). “Constraints on the Magnitude and Character of Host-rock Contamination.” Geochimica et Cosmochimica Acta, 74(14), 3733-3749.
-
Lippmann, J. et al. (2003). “Dating Ultra-Deep Mine Waters with Noble Gases and 36Cl, Witwatersrand Basin, South Africa.” Geochimica et Cosmochimica Acta, 67(23), 4597-4619.
-
Lin, L. H. et al. (2006). “Planktonic Microbial Communities from Underground Mines.” Geomicrobiology Journal, 23(6), 343-353.
-
Moser, D. P. et al. (2005). “Desulfotomaculum and Geobacter spp. in Ancient Saline Groundwater.” Geomicrobiology Journal, 22(7-8), 349-361.
-
Gold, T. (1992). “The Deep, Hot Biosphere.” Proceedings of the National Academy of Sciences, 89(13), 6045-6049.
-
Pedersen, K. (2000). “Exploration of Deep Intraterrestrial Microbial Life.” Science Progress, 83(1), 15-34.
-
D’Hondt, S. et al. (2004). “Distributions of Microbial Activities in Deep Subseafloor Sediments.” Science, 306(5705), 2216-2221.
-
Jørgensen, B. B. & D’Hondt, S. (2006). “A Starving Majority Deep Inside the Seafloor.” Science, 314(5801), 932-934.
-
Roussel, E. G. et al. (2008). “Archaeal Diversity and Distributions in the Deep Subsurface.” Environmental Microbiology, 10(3), 669-681.
-
Orsi, W. D. et al. (2013). “Life in the Ocean Subsurface.” Environmental Microbiology, 15(5), 1275-1284.
-
Morono, Y. et al. (2020). “Aerobic Microbial Life at 101.5 Million Years Old.” Nature Communications, 11(1), 3626.
-
Lloyd, K. G. et al. (2018). “Physiological Controls on Intraterrestrial Life.” Geobiology, 16(3), 241-248.
-
Kieft, T. L. et al. (2018). “Diversity of Life in Deep Subsurface Environments.” Frontiers in Microbiology, 9, 2852.
-
Starnawski, P. et al. (2017). “Environmental Comparisons Help Explain Disparate Microbial Communities.” Proceedings of the National Academy of Sciences, 114(12), 3154-3159.
-
Lever, M. A. et al. (2015). “Evidence for Microbial Carbon and Sulfur Cycling in Deeply Buried Ridge Flank Basalt.” Science, 348(6236), 709-713.
-
Lang, S. Q. et al. (2018). “From Subsurface Life to Mars: Building an Astrobiological Perspective.” Astrobiology, 18(7), 811-818.
-
Bradley, J. A. et al. (2020). “Widespread Energy Limitation to Life in Deep Subsurface Sediments.” Science Advances, 6(32), eaba0697.
-
Probst, A. J. et al. (2017). “Biomass Estimates for Deep Terrestrial Subsurface.” FEMS Microbiology Reviews, 41(5), 718-741.
-
Heberling, C. L. et al. (2014). “The Oxidation State of Deep Subsurface Crust.” Geochimica et Cosmochimica Acta, 131, 82-106.
-
Loman, N. J. et al. (2013). “A Robust Approach to Estimate Relative Microbial Biomass.” Nature Methods, 10(2), 99-102.